
Процедура Forever Young BBL способна омолаживать клетки кожи на генном уровне, делая их функционально подобными клеткам молодой кожи.
Anne Lynn S. Chang[1], Patrick H. Bitter Jr[2], KunQu Meihong Lin[1], and Howard Y. Chang [1, 3]
- Отделение дерматологии Стэнфордского университета, медицинский факультет, Redwood City, Калифорния, США;
- Advanced Aesthetic Dermatology, Los Gatos, Калифорния, США;
- Howard Hughes Medical Institute, Stanford, California, USA
Исследования на моделях организмов предполагают, что стареющие клетки могут быть функционально омоложены, но применима ли эта концепция к коже человека до сих пор оставалось недоказанным. В данном исследовании мы изучали экспрессию последовательности 30-концевых генов, чтобы исследовать изменения генов, связанных с фотостарением и естественным старением (хроностарением) кожи человека (собирательное понятие «старение кожи»), а также влияние процедур широкополосного света (BBL) на эти процессы. Мы считаем, что старение кожи ассоциировано со значительными изменениями в 2265 кодируемых и некодируемых РНК, 1293 из которых «омолаживались» после процедуры BBL; т.е. они становились более сходными по экспрессии генов с клетками молодой кожи. Омоложенные гены включали несколько уже известных ключевых регуляторов долголетия организмов в проксимальных некодируемых РНК. Старение кожи не связано с систематическими изменениями в кодировании 30-концевых мРНК (матричной РНК). Таким образом, процедура BBL может восстанавливать экспрессию генов в клетках с фотостарением и хронологическим старением кожи человека, подобно клеткам молодой кожи. Кроме того, наше исследование приводит новые аспекты, ранее не изученные другими авторами, которые позволяют по-новому взглянуть на процессы старения кожи человека.
Journal of Investigative Dermatology (2013) 133, 394–402; doi:10.1038/jid.2012.287; published online 30 August 2012
ВВЕДЕНИЕ
Старение является сложным процессом комплексных генетических изменений и воздействия окружающей среды. Старение связано с множественными изменениями экспрессии генов, каким образом изменения на генном уровне сказываются на здоровье человека до сих пор остается неизвестным. Многочисленные мутации единичных генов показывают продление жизни в моделях организмов (Partridge, 2010; de Magalhaes и соавт., 2012), ограничения в диете также могут замедлять скорость старения, даже если начинают использоваться в конце жизни (Partridge, 2010). Совсем недавно, в нескольких выступлениях было показано омоложение стареющих клеток или тканей, демонстрирующих удивительную обратимость процессов старения. Например, при гетерохронном парабиозе молодых и старых мышей высвобождаются циркуляторные факторы, которые восстанавливают функции стареющих стволовых клеток мышц (Liu and Rando, 2011). Кроме того, блокирование фактора транскрипции NF-kB в старых клетках эпидермиса мышей может прерывать клеточное старение и восстанавливать общую экспрессию генов клеток старой кожи, делая их подобными молодым клеткам (Adler et al., 2007). Возникает вопрос, обладают ли такой же обратимостью процессы старения в коже человека, где процессы старения развиваются в течение десятилетий, а не в течение месяцев или лет, как это было показано на моделях мышей. Выявление применимых в жизни методов блокирования процессов старения является серьезным вызовом.
Широкополосный свет (BBL), известный также как широкополосный импульсный свет - общедоступная и популярная процедура «омоложения» кожи, является идеальной технологией для тестирования этой концепции. Согласно данным Американского Общества Эстетической и Пластической Хирургии (ASAPS), более 215 миллионов Долларов потрачено на эти процедуры в 2009 году в США. В отличие от абляционных процедур, которые улучшают состояние кожи за счет термической деструкции и регенерации эпидермиса и поверхностного слоя дермы, BBL использует широкополосный некогерентный свет в диапазоне от 560 до 1200 нм, который поглощается многими компонентами кожи. В настоящее время процедуры BBL используются для уменьшения выраженности морщин, при нарушении пигментации, при эритеме и эластозе (Bitter Jr, 2000; Negishi et al., 2001). Тем не менее, молекулярные изменения, вызываемые этой процедурой не описаны.
«Омоложение» - термин, используемый как исследователями, так и широкими слоями населения, в который вкладывают различные понятия и, поэтому требуется определиться с понятием. В данном исследовании мы вкладываем в понятие «омоложение» восстановление показателей молодости в старых клетках и тканях. Действительно ли после процедуры BBL кожа омолаживается на молекулярном уровне и становится подобной молодой коже или же изменения связаны с раневыми процессами и регенерацией повреждений, что фундаментально отличается от молодой неповрежденной кожи?
Гистологически BBL снижает содержание меланина в дерме и устраняет телеангиэктазии (Bitter Jr, 2000; Prieton соавт., 2002), также в нескольких исследованиях сообщалось об увеличении содержания коллагена в верхних слоях сосочковой дермы к 3 неделе после процедуры (Negishi et al., 2001). Однако, синтез нового коллагена может носить изменчивый или краткосрочный эффект, согласно данным ультраструктурного анализа кожи через 3 месяца после процедуры не показало признаков влияния на коллаген и эластин (Prieto et al., 2002). Мы изучили молекулярные основы ответа на процедуры BBL, определяя общую экспрессию генов кожи человека с фотостарением и хронологическим старением после процедур BBL. Целью являлось отследить широкий спектр изменений в РНК стареющих клеток при воздействии BBL, включая изменения экспрессии генов (кодируемых и некодируемых) и регуляцию генов.
РЕЗУЛЬТАТЫ
Клинические и гистологические изменения после процедуры BBL
Чтобы получить представление о экспрессии генов связанных со старением кожи и процедурой BBL мы использовали биопсию кожи молодых женщин волонтеров (возраст менее 30 лет, n=5) и биопсию участков стареющей кожи женщин без каких либо воздействий (контроль) и участков стареющей кожи женщин-добровольцев после процедуры BBL (возраст старше 50 лет, N=5), после трех курсов процедур BBLодин раз в месяц (n=5, Рисунок 1а). Процедуры проводились на здоровых пожилых женщинах с умеренным и выраженным фотоповреждением кожи предплечий, проживающих в Санта Клара или Сан Хосе, Калифорния, где 257 солнечных дней из 365 дней со средним UV-индексом 5,1 (средний UV-индекс в США 4,3; источник: www.bestplaces.net, данные 25 апреля 2012 г.). Солярий, местное нанесение ретиноиды и любые другие процедуры на коже рук были запрещены за 1 месяц до начала исследования и в течение исследования. На протяжении всего исследования участники были проинструктированы защищать свои руки с помощью солнцезащитных кремов широкого спектра и одежды с длинными рукавами, а также избегать лишнего контакта с солнцем. Молодые участницы исследования имели те же критерии включения в исследование, но не имели признаков фотостарения на руках.
После трех процедур BBL, на коже рук происходили улучшения клинических показателей хронологического старения и старения кожи под воздействием факторов внешней среды: мелкие морщинки (Р=0.03), пигментация (Р=0.02) и общие показатели старения кожи (Р=0.01; Рисунок 1а-с). По данным гистологического исследования, признаки эластоза в образцах стареющих клеток с участков, подвергшихся процедуре BBL, были менее выражены, в сравнении с образцами стареющей кожи, не подвергавшихся воздействию BBL (Рисунок 1d-g). Окрашивание гистологических образцов методом PAS не показало изменений в количестве коллагена в дерме между леченными и не леченными образцами кожи, хотя после процедуры BBLволокна коллагена были менее дезагрегированы (Рисунок 1h и i). Образцы стареющей кожи после процедуры показывали субъективное увеличение плотности эпидермиса (Рисунок 1e, g, i) в сравнении с не леченными образцами стареющей кожи (Рисунок 1d, f, h).
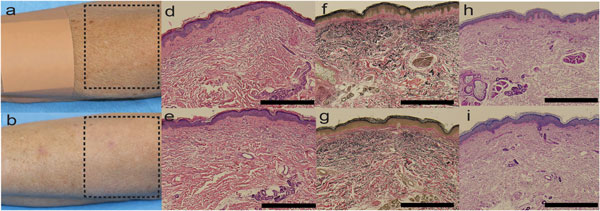 Рисунок 1. Клинические и гистологические эффекты, фотоомоложение BBL.
Рисунок 1. Клинические и гистологические эффекты, фотоомоложение BBL.
(а) Рука женщины 73-лет до процедуры BBL (штрих-линия указывает зону процедуры и повязка указывает контрольную зону, не подвергающуюся лечению). (b) та же рука после трех процедур BBL: отмечается редукция мелких морщин, уменьшение гиперпигментации и эритемы на обработанном участке кожи (зона, обозначенная штрих-линией), в сравнении с необработанным участком. (с) Показатели старения кожи – значительное уменьшение выраженности мелких морщин, уменьшение выраженности пигментации и снижение общего показателя старения кожи после процедуры BBL. Значение Р. Согласно двухстороннему t-тесту. (d) Гистология кожи до процедуры BBL показывает наличие эластоза (увеличение х200, окрашивание гематоксилин-эозин (ГЭ)) и (е) редукция эластоза (х200, ГЭ) после процедуры BBL. (f) До процедуры, явный эластоз (увеличение х200, окрашивает по von Giesen). (g) После процедур эластоз менее выражен (увеличение х200, окрашивает по von Giesen). (h) До процедуры волокна коллагена дезорганизованы и неупорядочены (увеличение х200, окрашивание PAS). (i) После процедур волокна более однородные (увеличение х200, окрашивание PAS). Цена деления 1 мм.
| Клинические показатели | Без лечения (n=5), среднее значение (SD) | После процедуры (n=5), среднее значение (SD) | P |
|---|---|---|---|
| Мелкие морщинки | 3,2 (1,3) | 1,0 (1,0) | 0,02 |
| Выраженные морщины | 0 (0) | 1,8 (2,5) | 0,18 |
| Повышенная пигментация | 7,2 (1,3) | 3,4 (2,3) | 0,02 |
| Общий результат | 6,6 (1,1) | 3,4 (1,5) | 0,01 |
Экспрессия кодирующих и некодирующих РНК в стареющей коже
Хотя экспрессия генов стареющих клеток в некоторых тканях была изучена ранее методом гибридизации микромассивов, мы использовали 3'-концевую последовательность для количественной оценки экспрессии (3-seq), как эффективный метод оценки глубоких 3'-концевых последовательностей РНК (Tariq et al., 2011). Потенциальные преимущества 3'-концевых последовательностей дают возможность точного определения уровней транскрипции без затемнения кросс-гибридизацией, а также возможность определить повреждения в окончаниях РНК и процессинга, а также выявить не описанные ранее гены, таких как длинные некодируемые РНК (IncRNA). Мы создали 6,5-12,4 миллиона уникальных картирований для каждого образца и идентифицировали участки генов с различием экспрессии при помощи DESeq алгоритма (смотри Материалы и методы).
Чтобы точно выявить признаки старения в окончаниях молекул, мы вначале выявляли нарушения транскрипции участков генов, ассоциированных со старением путем сравнения образцов молодой кожи с образцами не леченной стареющей кожи, а затем тестировали, какое влияние оказывает процедура BBL на изменение этих показателей.
Сравнение уровней транскрипции мРНК (матричная РНК) в не леченной молодых в сравнении с не леченной стареющими, а также стареющих не леченных в сравнении со стареющими леченными, образцами показало значительное увеличение уровней экспрессии в 3530 генах (Рисунок 2а). Направленность изменения генов после процедуры BBL показано на Рисунке 2а, голубым отмечено 2-кратное уменьшение и желтым 2-кратное увеличение. Гены, у которых уровень транскрипции не леченных молодых и не леченных стареющих образцов претерпел существенные изменения опубликован онлайн в дополнительных материалах к исследованию в таблице S1 (доступен по адресу оригинальной онлайн публикации JID).
Для наглядного визуального отображения значимых генов мы использовали визуальную тепловую карту (Рисунок 2а), справа от которой расположили таблицу, пурпурные полосы которой отображают основные биологические эффекты, за которые отвечают гены, в соответствии с генной онтологией. Например, «омоложенные гены» и lncRNAs (длинные некодируемы РНК) расположены в обоих верхних и нижних частях визуальной тепловой карты. Гены, отвечающие за иммунный ответ и трансляцию, расположены в нижней половине карты. Гены, отвечающие за клеточную адгезию, расположены в верхней части карты. Активность этих генов снижена в группе молодых волонтеров и увеличена в группе не леченных волонтеров со стареющей кожей, а в группе леченных волонтеров со стареющей кожей активность генов адгезии находится на среднем уровне. Соседняя таблица с пурпурными столбцам указывает на то, в каких генах происходят изменения и за какие биологические процессы отвечают эти гены. Визуальна тепловая таблица при помощи «теплых» (желтый, повышение) и «холодных» (синий, снижение) цветов указывают направленность изменения активности генов - увеличение или снижение их функции, что позволяет сравнивать молодую кожу, стареющую нелеченную и стареющую леченную кожу. Например, молодая кожа и леченная старая кожа показывают увеличение уровней транскрипции, отвечающих за «иммунный ответ» и «трансляцию», поскольку обе эти группы показали увеличение активности генов (желтый цвет). И наоборот, в группе не леченных пациентов со стареющей кожей происходит снижение активности генов (синий) «иммунного ответа» и «трансляции» в сравнении с двумя другими группами.
Генные изменения, связанные со старением, многогранны и объединяют в себе изменения нескольких биологических функций. Первая пятерка наиболее выраженных изменений с повышением активности генов стареющей не леченной кожи, в сравнении с молодой не леченной кожей включает: трансляция (Р=4,7х10-12), трансляционное удлинение (Р=5,1х10-7), макромолекулярные сложные комплексы (Р=7,5х10-6), нкРНК (не кодируемые РНК) отвечающие за метаболизм (Р=6,2х10-6) и РНК процессинг (Р=2,5х10-6). Первая пятерка генов со снижением функции в группе стареющей не леченной коже в сравнении с молодой кожей включала: гены, отвечающие за клеточную адгезию (Р=1,5х10-17), биологическая адгезия(Р=1,5х10-17), гомофильная клеточная адгезия (Р=7,8х10-8), ген, отвечающий за развитие скелета (Р=3,2х10-7), энзим-связанный белок-рецептор сигнального пути (Р=5,2х10-6). Изменения в этих генах напоминают изменения набора генов при старении в других тканях и организмах. Например, гены, связанные с трансляцией или регуляторы трансляции связаны со старением Caenorhabditis elegans(Long et al., 2002) и Drosophila melanogaster (Kirby et al., 2002). Кроме того, как полагают, трансляция играет важную роль вTOR(мишень для рапамицина) процессе старении стволовых клеток (Chen и соавт., 2009; Nelson и соавт., 2009; Liu и Rando, 2011; Serrano, 2011).
BBL способствует изменению экспрессии генов, делая их подобными молодым клеткам кожи
Гены, уровень экспрессии которых в стареющей коже, леченной BBL, был ближе к генам молодой не леченной кожи, были обозначены RGs. Это означает, что уровень экспрессии генов группы стареющей леченной кожи был вычтен из уровней экспрессии генов не леченной молодой кожи, также как из не леченной стареющей кожи. Если разница в уровне экспрессии генов была ниже в сравнении с молодой кожей, в сравнении разницей с не леченной стареющей кожей, то данный ген считался «омоложенным». Суммарно 1293 транскриптов генов квалифицировали, как омоложенные гены, RGs (Детально описано в дополнительных таблицах S2). Иерархическая кластеризация показала, что экспрессия генов леченной стареющей кожи более сходна с не леченной молодой кожей, чем с не леченной стареющей кожей (Рисунок 2а). Омоложенные гены отвечают за сцепленные с ними определенные биологические функции, шесть наиболее выраженных из них: трансляция (P=5,8х10-11), РНК процессинг (Р=6,3х10-8), нкРНК процессинга метаболизма (Р=1,4х10-7),регуляция клеточного метаболизма белка (Р=1,6х10-5), макромолекулярные катаболические процессы (Р=2,1х10-5),клеточный цикл (Р=2,4х10-5; (Рисунок 2b, справа вверху).
Более детальное исследование экспрессии «омоложенных» процедурой BBL генов связанных с ключевыми регуляторами, известными, как гены, контролирующие старение (Рисунок 2с). Они включают ZMPSTE24, металлопротеиназу, которая перерабатывает ламин А, дефект этого гена вызывает синдром выраженного преждевременного старения, прогерию Хатчинсона-Гилфорда. Кроме того, рецептор IGF1R был одним из «омоложенных генов», этот ген непосредственно связан со старением и долгожительством в моделях человека, мышей и других организмов (Liang et al., 2011; Tazearslan et al., 2011). Другие «омоложенные гены» EIF4G1 и EIF4EBP1 отвечают за повышение продолжительности жизни у C. elegans (Curran и Ruvkun, 2007). MLL– регулятор транскрипции, связанный с теломерами (Caslini et al., 2009), и метилат H3K4, который отвечает за нормальную продолжительность жизни у C.elegans (Greer и соавт., 2010). MAP3K5 (ASK10) регулирует активность киназ в ответ на оксидативный стресс в модели стареющих мышей Klotho (Hsieh и соавт., 2010). PSMD8 является компонент протеасом, нарушение работы которых способствует старению кожи человека (Hwang и соавт., 2007). RING1 и MOV10 участвуют в процессах, которые контролируют длительность жизни фибробластов человека (Itahana и соавт., 2003). EEF2 (эукариотический фактор трансляции элонгации 2) был также отнесен к «омоложенным генам», согласно исследованиям, связан со снижением синтеза белка у крыс (Parado и соавт., 1999). Наконец, ряд генов-супрессоров, которые контролируют клеточный цикл и обеспечивают целостность генома, таких как ING4 супрессор опухоли, DAXXи MSH2, были также «омоложены». Таким образом, процедура BBL способна восстанавливать множество молекулярных свойств молодой кожи в стареющих клетках кожи человека, по меньшей мере, в краткосрочной перспективе. К тому же, мы не обнаружили изменений экспрессии генов, связанных с раневыми процессами и процессами рубцеобразования.
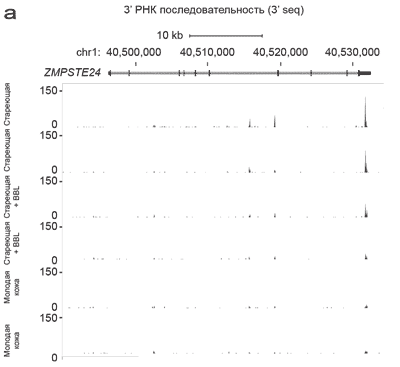
С целью подтверждения наших находок методом 3-seq анализа, мы провели количественную ПЦР с обратной транскрипцией (qRT–PCR), чтобы проверить уровни ZMPSTE24 в независимых группах женщин в разном возрасте. 3-seq анализ показал, что не леченная стареющая кожа имела наивысшие уровни экспрессии транскрипции ZMPSTE24, леченная стареющая кожа имела средние уровни экспрессии ZMPSTE24, а не леченная молодая кожа имела самые низкие уровни (Рисунок 3а). Методом обратной ПЦР реакции определили, что стареющая кожа предплечья (возраст 75 лет) имела высокие уровни транскрипцииZMPSTE24, не леченная кожа предплечья в среднем возрасте (возраст 35 лет) имела средние уровни и не леченная молодая кожа предплечья (возраст 24 года) имела самые низкие уровни (Рисунок 3b). Такое соотношение не было описано ранее для человека, но корреляция двух независимых методах дает основание предполагать, что наши находки свидетельствуют о взаимосвязи биологических процессов с процессами физиологического старения.
Увеличение генов, кодирующих мРНК, участвующих в РНК процессинге, побудило нас исследовать уровни экспрессии дополнительных классов РНК. Длинноцепочечная некодируемая РНК (lncRNA) относится к новому классу генетических элементов, которые широко транскрибируются в геноме человека (Wang и соавт., 2009; Wapinski и Chang, 2011). Роль lncРНК в процессах старения и ее уровни в коже не были изучены, так как ранее они не были картированы. Тем не менее, 3-seq методика может легко идентифицировать и количественно определять экспрессию lncRNA. Из 3530 транскриптов с измененными уровнями между не леченной молодой и не леченной стареющей кожей, 151 были lncРНК. Расположение в хромосомах и более детальная информация о генах этих lncRNA приведена в дополнительной таблице S3 онлайн. Из 1293 омоложенных генов, 42 были lncРНК. в хромосомах и более детальная информация о генах этих lncРНК приведена в дополнительной таблице S4 онлайн с тепловой картой. Эти находки свидетельствуют, что lncРНК потенциально вовлечены в процессы старения и омоложения, параллельно их роли в развитии и клеточном перепрограммировании (Gupta и соавт., 2010; Loewer и соавт., 2010). Наши данные являются первым сообщением о связи lncРНК со старением человека, что может послужить основой для более детальных исследований в будущем.
Генетический анализ 151 lncРНК с выраженными отличиями экспрессии между молодой не леченной и стареющей не леченной кожей не выявил существенных различий. Генетический анализ 42 «омоложенных» lncРНК также не показал существенных изменений. Тем не менее, это не уменьшает значимость lncРНК в «омоложении». Например, небольшое количество из 42 lncРНК дает основания полагать, что дальнейшие исследования помогут выявить другие lncРНК с выраженными биологическими эффектами. К тому же, lncРНК являются новым классом РНК и наш анализ позволит приблизить понимание роли lncРНК последовательностей в изучении генов; возможно, что lncРНК играют важную роль в регуляции генов, удаленных от расположения lncРНК (Gupta и соавт., 2010).
Влияние процедуры BBL на иммунный ответ отражается в изменении показателей иммунного ответа, как в образцах не леченной молодой кожи. Рисунок 2а показывает, что хотя гены, отвечающие за иммунный ответ «повышают» активность после процедуры, этот профиль «повышения» больше напоминает молодую не леченную кожу. Это предполагает, что небольшая часть иммунного ответа, которая «повысилась» после процедуры является частью «омоложения» и является неспецифическим ответом на процедуру BBL.
В исследованиях было показано, что NF-kB путь является важным в старении кожи и омоложении (Adler et al., 2007), и мы обнаружили, что «омоложенные гены», связанные с NF-kB становятся высоко активными. В целом, 827 из 1293 генов связаны с NF-kB (Р=1,2х10-75, гипергеометрический тест). Интересно, что непосредственно ген NF-kB не вошел в число «омоложенных генов».
3' окончания РНК выглядят неповрежденными при старении или после BBL
3-seq выборки 3' полиаденилированных РНК (полиА) фрагментов для более глубокого исследования, и следовательно, имеют потенциал для выявления изменений, локализующихся в 3' транскрипт-окончаниях. Метод 3'-seq последовательностей образцов РНК, идущих вверх от полиаденилированный окончаний. Если на этих участках имеются признаки изменений полиаденилированных участков в пределах последнего экзона, то последний экзон удлиняется или укорачивается, что может быть замечено в тестах. Этот метод не позволяет исследовать длину полиаденилированных окончаний.
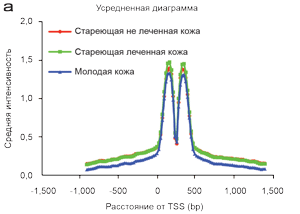
Альтернативное использование 3'-окончаний является важным регуляторным механизмом (Mayr и Bartel, 2009), и может изменить экспрессию генов путем изменения содержимого 3' нетранслируемого региона, что может оказать влияние на набор мишеней микроРНК или РНК-связывающих протеинов (как те, что наблюдаются при раке, Shapiro и соавт., 2011). Таким образом, дополнительно к количественным изменениям транскрипционных множеств, мы также изучали изменения в окончаниях транскриптов, связанных со старением и процедурами BBL. Системное сравнение всех 3-seq образцов показало, что, как и предполагалось, большинство считываний попало на упомянутый последний экзон, то есть <1000 bp от стоп-кодона транскрипции (Рисунок 3а), и не было соответствующих изменений 3'-окончаний, связанных с возрастом или процедурой BBL (Рисунок 3b). Например, если распределение расстояний от стоп-кодона транскрипции для РНК молодых не леченных и стареющих леченных образцов были различны, то старение может быть связано с системными изменениями в мРНК (матричная РНК) 3' окончаниях (Рисунок 4).
|
|
|
|
|
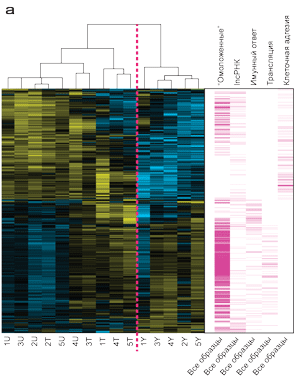
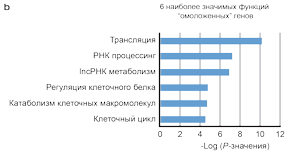

Рисунок 2. Эффект широкополосного света BBL на кодируемые и некодируемые РНК в стареющей коже.
(а) Кластеризация экспрессии генов леченной стареющей кожи расположена посредине между не леченной молодой и не леченной стареющей кожей. Показаны уровни транскрипции, которые претерпели значительные изменения у не леченой молодой в сравнении с не леченной стареющей кожей, а также не леченная стареющая кожа в сравнении со стареющей леченной кожей (n=3530 транскриптов). Столбцы соответствуют образцам (группам исследования), ряды соответствуют отдельным генам. Т – стареющая леченная кожа, U – стареющая не леченная кожа, Y – молодая не леченная кожа. Столбцы пурпурного цвета отображают распределение генов на большой «тепловой» карте (тепловая карта – способ визуального отображения выраженности изменений, где холодные тона указывают на снижение функции, а теплые тона на степень увеличения функции), сгруппированных по биологическим функциям. Например, «иммунный ответ» и «трансляция» соответствующих генов расположены на нижней половине тепловой карты, желтые ряды указывают на повышение экспрессии в группах леченной стареющей кожи и не леченной молодой группы; соответственно, расположение на тепловой карте для иммунного ответа и трансляции обозначено голубым (т.е. функция снижена) в группе не леченной стареющей кожи. Распределения на тепловой карте «омоложенных» генов (n=1293) и «длинных некодируемых РНК» (lncРНК) показано в первом и втором пурпурных столбцах, соответственно. (b) Шесть наиболее значимых функций «омоложенных» генов. (с) Образцы омоложенных генов с известной антивозрастной функцией.
 |
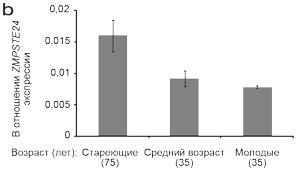 |
Рисунок 3. Уровень транскрипции ZMPSTE24 снизился после процедур BBL.
(а) схематическое отображение локуса ZMPSTE24 на хромосоме 1, hg18. Уровень транскрипции ZMPSTE24 снизился после процедуры BBL. (b) Экспрессия ZMPSTE24 транскрипции выше в не леченной стареющей коже в сравнении с не леченной молодой кожей, определено методом обратной ПЦР.
 |
 |
Рисунок 4. Процедура BBL при старении не оказывает влияния на обработку 3' окончаний.
(a) системное сравнение всех 3-seq (расширенное исследование последовательностей РНК 3' окончаний) показывает, что большинство считываний приходится на последний упомянутый последний экзон (расположенный на расстоянии 1,000 bp от участка начала транскрипции (TSS)) для не леченных стареющих, леченных стареющих и не леченных молодых. Ось Y показывает среднюю интенсивность 3-seq сигнала. (b) Не выявлено систематических изменений в использовании 3’-окончаний связанных с процедурой BBL, поскольку считывания показали одинаковые длины распределений у не леченных стареющих, леченных стареющих и не леченных молодых.
Специфический ответ на процедуру BBL
В дополнение к эффектам, связанным с экспрессией возраст-ассоциированных генов, мы также рассмотрели возможность того, что процедура BBL может оказывать специфическое лечебное действие, не связанное со старением. Например, что процедура BBL могла бы стимулировать процессы заживления ран или рубцеоразования, в дополнение к омолаживающему действию. Мы выявили существенные изменения экспрессии 1112 генов, которые наблюдались только в образцах прошедших процедуру BBL, но этих изменений не было выявлено в образцах не леченной молодой и не леченной стареющей кожи. Среди этих генов со специфическим ответом на процедуру, пять наиболее выраженных биологических эффектов с увеличением экспрессии генов после процедуры были: иммунный ответ (Р=3,8х10-12), положительная регуляция иммунной системы (Р=2,0х10-8), активация клеток (Р=5,7х10-8), активации Т-клеток (Р=6,0х10-7) и защитные реакции (Р=1,4х10-7). Эти реакции связаны с иммунным ответом на BBL и отличаются от иммунного ответа генов, которые также увеличивались у не леченых молодых (как указывалось выше). Пять наиболее значимых биологических эффекта, связанных со снижением экспрессии генов после процедуры были: регуляция транскрипции (Р=2,0х10-6), транскрипция (Р=1,7х10-5), ответ на органические вещества (Р=1,1х10-4), ответ на гормональные стимулы (Р=4,4х10-4) и отрицательная регуляция транскрипции (Р=4,7х10-4). Эти гены отличаются от предварительно описанных эффектов на «заживление», которые характеризуют ответ процессы заживления кожи (Chang и соавт., 2005); однако сложно сделать прямое сравнение, поскольку нет описанных в литературе эквивалентных нашим данным по активности генов, связанных раневыми процессами на 4-ю неделю.
И наконец, топ 10 генов, повышающих активность и снижающих активность у леченных BBL со стареющей кожей в сравнении с не леченной стареющей кожей приведены в Таблица 1.
Таблица 1. Топ 10 генов, повышающих активность и снижающих активность у леченных BBL со стареющей кожей в сравнении с не леченной стареющей кожей.
| Обозначение гена | Направленность изменений: леченная стареющая кожа в сравнении с не леченной стареющей кожей | Направленность изменения активности |
|---|---|---|
| HEPHL1 | 3,19 | ↓ |
| ZNF660 | 3,09 | ↓ |
| LY6G6D | 2,53 | ↓ |
| COCH | 2,38 | ↓ |
| CCL18 | 2,36 | ↑ |
| CEP78 | 2,36 | ↓ |
| ANGPTL7 | 2,34 | ↓ |
| SLN | 2,17 | ↓ |
| CPXM1 | 2,10 | ↑ |
| SAMD5 | 2,08 | ↓ |
| Аббревиатура: BBL Broadband light, широкополосный свет. | ||
ДИСКУССИЯ
Полученные нами результаты показывают, регуляторы старения организма могут быть изменены в коже человека благодаря использованию общедоступной технологии BBL. Насколько пластичны процессы старения и насколько они могут изменяться в сторону оздоровления кожи, таких как профилактика или лечение ассоциированных со старением состояний кожи еще предстоит выяснить. Хотя технология BBL используется больше для придания коже внешне «молодого» вида, наше исследование показало, что наблюдается омоложение кожи на молекулярном уровне, с многочисленными изменениями в работе генов, связанных с процессами старения, изменения экспрессии генов стареющей кожи становится подобной генам молодой кожи. Таким образом, что видимые клинические изменения обусловлены функциональным омоложением кожи (как минимум на короткий период времени), а не внешней косметической имитацией вида молодой кожи.
Поскольку технология BBL существует менее 20 лет, долгосрочные эффекты BBL терапии еще предстоит определить. Хотя в данном исследовании оценивали состояние кожи через 4 недели после процедуры, неясно, как долго сохраняется этот клинический эффект и молекулярные изменения. Неизвестно также, насколько стойко сохраняется эффект у таких связанных с возрастом состояний, как себорейный кератоз или актинический кератоз со временем. Может быть информативным исследовать нынешних участников в долгосрочной перспективе (45 лет) с фотографированием и биопсией кожи, чтобы определить длительность клинического, гистологического и молекулярного эффекта процедуры BBL.
Точные механизмы, при помощи которых BBL (некогерентное световое излучение) изменяет экспрессию генов, на данный момент хорошо не изучены. Например, известно, каким образом BBL поглощается различными мишенями – меланином и гемоглобином, что приводит к уменьшению эритемы и пигментации. Считают, что уменьшение выраженности мелких морщинок частично связано с синтезом нового коллагена (Fisher и соавт., 2008). В на протяжении данного исследования не было выявлено влияния на гены, связанные обменом коллагена. Возможно, что если бы биопсия проводилась в более ранние сроки, чем 4 недели, некоторые изменения экспрессии генов, связанные с синтезом коллагена, могли бы быть зафиксированы изменения.
Экспрессия генов, связанная со старением человека различна для различных органов. Например, старение почек и мышц человека имеют изменения в других генах (Rodwell и соавт., 2004; Zahn и соавт., 2006). Данные, полученные для старения кожи в данном исследовании отличаются от генетических данных старения других органов; однако последующие сравнения могут пролить свет на процессы старения в целом.
NF-kB является важным регулятором экспрессии генов в различных контекстах. В данном случае, наиболее релевантные свойства NF-kB заключаются в контроле клеточного старения (Bernard и соавт., 2004; Adler и соавт., 2007) и иммунного ответа. BBLможет оказывать влияние на механизмы, контролируемые NF-kB, на что указывает высокое содержание «омоложенных» NF-kB-связанных генов. Точные механизмы, которые задействованы в этих процессах подлежат дальнейшему изучению. Тем не менее, полученными нами результаты соответствуют предыдущим исследованиям, показавшим, что индуцируемая блокада NF-kB в коже стареющих мышей, восстанавливает экспрессию генов и фенотип молодой кожи (Adler и соавт., 2007).
Сложно напрямую сравнить результаты нашего исследования с профилями экспрессии генов у людей, описанными в настоящее время в литературе по двум причинам: (1) различием во времени биопсии и (2) причина болезни или ее лечения отличаются от процедуры BBL. Например, экспрессия генов в пост ожоговом гипертрофичеком рубце у человека к 6-15 месяцам у двух детей и двух взрослых приводит к существенной активации шести генов (Paddock и соавт., 2003), эти гены не активировались после процедуры BBL в нашем исследовании. Другой пример, в модели царапины на кератиноцитах человека в in vitro происходит активация NF-kB в клетках между 1 и 14 днями (Adams и соавт., 2007). В нашем исследовании тестирование проводилось через 1 месяц после процедуры, когда эффект реакции репарации раны мог снижаться и мы зафиксировали преимущественно омолаживающий эффект. К моменту тестовой точки в 1 месяц уровни NF-kB не были существенно увеличены, но активность генов, взаимодействующих с NF-kB, была существенно увеличена, что является явным и, насколько нам известно, ранее никем не описано.
Два «омоложенных» гена RING1 и MOV10 Polycomb пути с потенциальным эффектом как на заживление раны, так и на омоложение. У мышей и в клеточных культурах, Polycomb путь контролирует продолжительность жизни фибробластов человека (Itahana и соавт., 2003) и ассоциируется с повышением активности генов, отвечающих за процессы заживления ран (Shaw и Martin, 2009).
Лиганды к Толл-подобным рецепторам 2, 3 и 5, как сообщалось влияют на транскрипцию и уровень протеинов металлопротеиназ 1 и 9 матрикса и запускало ядерную транслокацию NF-kB после 24-48 часов в культуре кератиноцитов человека (Lee и соавт., 2009). Мы не отметили существенного повышения Толл-подобных рецепторов 2, 3 и 5 или NF-kB, но наше исследование было in vivo и образцы кожи были получены к концу 1 месяца.
К тому же, наши данные показывают, что сцепленные биологические эффекты, такие как «трансляция» или «РНК процессинг» изменялись после процедуры BBL, наши исследования не выявляли популяций клеток кожи, в которых происходили эти изменения. Следующие исследования, возможно, позволят объяснить, каким образом процедура BBL может приводить к гистологическим или структурным изменениям, таким как резорбция эластоза или скоплений коллагена.
Было бы интересно сравнить другие факторы, известные своими клиническими результатами воздействия на старение кожи, такими как используемый местно третиноин, подобный по изменениям экспрессии генов изменениям, наблюдаемым в генах после процедуры BBL. Кроме того, сравнение участков стареющей кожи, не подвергающихся воздействию солнца, позволит идентифицировать изменения в экспрессии генов, специфически связанных с хроностарением кожи. Это предварительное исследование и мы будем изучать влияние BBL на молодую кожу. В данном исследовании мы не проводили процедур BBL на молодой коже (у лиц в возрасте моложе 30 лет), поскольку они не имели клинических показаний; эти лица не имели различимых признаков фотостарения и хроностарения кожи предплечий. Поскольку маловероятно, что BBL будет использоваться на практике на молодой коже без признаков фотостарения (за исключением удаления волос), мы не включили эту группу в исследование.
Данные о возможном влиянии BBL на синтез коллагена противоречивы, согласно доступным источникам. Хотя некоторые доклады указывают на гистологические изменения, индуцированные процедурой BBL, включая неогенез коллагена (Negishi и соавт., 2001), в других исследованиях этих изменений не отмечается (Prieto и соавт., 2002). Согласно данным последнего исследования, не отмечено также изменений в содержании эластина после процедуры. В нашем исследовании не было отмечено существенных изменений в содержании коллагена после процедуры на окрашенных по PAS образцах. Они имели снижение количества эластина на препаратах, окрашенных по von Giesen. Мы не обнаружили существенных изменений уровней экспрессии генов коллагена и эластина после процедуры. Возможно, причина заключается в одном заборе материалов для тестов, что могло не захватить промежуток времени, когда уровень экспрессии генов коллаген и эластин претерпевал существенные изменения. Последующие исследования с биопсией кожи позволит выявить кинетику процессов активации / супрессии генов-мишеней. Кроме того, целью данного исследования было выйти за рамки простого гистологического анализа кожи и изучить молекулярные изменения при старении кожи и влияние на них процедур BBL. Мы исследовали множество изменений в генах соединительной ткани, которые могут модулироваться BBL.
Последующие исследования с большими выборками могут позволить нам выявить дополнительные значимые гены (как кодируемые, так и не кодируемые), экспрессия которых изменяется в сравнении стареющей не леченной кожи с молодой не леченной кожей, а также не леченной стареющей кожей с леченной стареющей кожей. Большие выборки могут позволить соотнести степень клинического ответа с изменениями в «омоложенных» генах.
МАТЕРИАЛЫ И МЕТОДЫ
Участники исследования и материалы
Данное исследование проведено в соответствии с Хельсинской декларацией. После ознакомления и подписания информированного согласия, пяти женщинам-участницам в возрасте старше 50 лет провели процедуры BBL на левом предплечье. Критериями включения в исследование были тип кожи II или IIIпо Фитцпатрику и умеренная или выраженная степень старения кожи на предплечье (McKenzie и соавт., 2010) для участников группы лечения. Процедура проводилась на платформе Joule компании Sciton с использованием модуля BBL. Участникам проводили 3 процедуры с интервалом 4 недели с использованием фильтров 515-нм или 560-нм одним импульсом, длительностью 10-20 мс с удельной мощностью 8-14 Дж/см². Во время одной процедуры выполнялось два или более прохода. На 4-ю неделю после третьей процедуры BBL проводилась Punch-биопсия участков кожи размером 4 мм на обработанной коже и соседнем необработанном участке кожи. Также проводилась Punch-биопсия (4 мм) с участков кожи, не подвергающихся воздействию солнечных лучей у 5 участниц возрастом до 30 лет. Эти образцы были разделены пополам и помещены в RNAlater (раствор для стабилизации РНК - Ambion, ном. По кат.AM7022,GrandIsland, NY) или раствор формалина для окрашивания гематоксилин-эозином, по von Giesen или PAS.
3-seq и биоинформатика
Все РНК экстрагировались при помощи RNeasy Fibrous Tissue Mini Kit (Qiagen, Germantown, MD). 3-seq методика проводилась согласно Beck и соавт. (2010). Суть данной методики вкратце заключается в следующем: oligo-dT-направленная обратная транскрипция генерирует комплиментарные ДНК, соответствующие 3' окончаниям полиА транскриптов; затем клонировались комплиментарные ДНК и подвергались глубокому анализу последовательностей на платформе Illumina GAIIx (San Diego, CA) с размером считывания материала 36 bp. Считываемые материалы были приведены в соответствие с геномом человека (hg18) при помощи bowtie (Langmead et al., 2009); каждый образец генерировал 6,5-12,4 миллиона уникальных картируемых считываний. 3’ последовательности транскриптов кожи исследовались на распределение по длине.
Считывание на тысячу оснований экзона на один миллион картируемых считываний (RPKM, прямое измерение множества последовательностей транскрипции) и многочисленные считывания материалов, приходящиеся на каждый ген были рассчитаны с использованием самостоятельно-разработанных сценариев Kun Qu. Соответствующие последовательности (RefSeq; http://www.ncbi.nlm.nih.gov/refseq/) и Ensembl (http://www.ensembl.org) описывающие некодируемые гены были также включены. Существенные гены были изучены с использованием набора DESeq (http://www.bioconductor.org) сравнением стареющей леченной со стареющей не леченной кожей (изменения генов, вызванные процедурой), и стареющие не леченные с молодыми не леченными образцами (изменения генов, вызванные старением). Неконтролируемая иерархическая кластеризация генов с существенно различающимися экспрессиями генов проводилась с использованием кластеров. Биологические маркеры генерировались с использованием DAVID (Database for Annotation, Visualization and Integrated Discovery) Bioinformatics Resources 6.7 (http://david.abcc.ncifcrf.gov/). Гены близкие к lnc-генам были идентифицированы при помощи базы данных GREAT (http://great.stanford.edu). Эти данные были помещены в базу данных Gene Expression Omnibus.
Для определения взаимосвязи между «омоложенными» генами и NF-kB связями, мы загрузили NF-kB-связанные гены идентифицированные проектом ENCODE (ENCODE Consortium, 2011) методом иммунопреципитации хроматина. Всего 9650 генов связаны с NF-kB в одном или более типах клеток, мы сравнивали их со списком «омоложенных» генов.
ПЦР с обратной транскрипцией
Все РНК были экстрагированы при помощи TRIzol (Invitrogen, Grand Island, NY) с последующей колоночной очисткой RNeasy (Qiagen) и DNAse Turbo Treatment (Ambion). ПЦР с обратной транскрипцией проводилась с использованием РНК в количестве 10 нг, на Taqman One Step RT–PCR master mix и на одной из следующих проб Taqman: GAPDH (Hs99999905_m1) и ZMPSTE24 (Hs00956778_m1; Applied Biosystems, Carlsbad, CA). Реакции проводились в тех экземплярах для каждого образца и минимум 2 раза. Данные были нормализированы по уровням глицеральдегид-3-фосфат дегидрогеназы (GAPDH).
КОНФЛИКТ ИНТЕРЕСОВ
PatrickBitter является докладчиком по технологиям широкополосного света. Другие авторы не имеют конфликтов интересов.
БЛАГОДАРНОСТИ
Данное исследование было проведено, благодаря исследовательскому грану от Sciton (Сайтон). Мы благодарны Paul Khavari and Jean Tang за подготовку материалов к данной публикации. Мы благодарим Елену Михайличенко и Sarah Jacobs за административную поддержку.
СПИСОК ЛИТЕРАТУРЫ
- Adams S, Pankow S, Werner S et al. (2007) Regulation of NFKB activity and keratinocytic differentiation by the RIP4 protein: implications for cutaneous wound repair. J Invest Dermatol 127:538–44
- Adler AS, Sinha S, Kawahara TLA et al. (2007) Motif module map reveals enforcement of aging by continual NFKB activity. Genes Dev 21:3244–57
- Beck AH, Weng Z, Witten DM et al. (2010) 30-end sequencing for expression quantification (3SEQ) from archival tumor samples. PLoS One 19:5: e8768
- Bernard D, Gosselin K, Monte D et al. (2004) Involvement of Rel/ NFkB transcription factors in keratinocyte senescence. Cancer Res 64:472–81
- Bitter Jr PH (2000) Noninvasive rejuvenation of photodamaged skin using serial, full-face intense pulsed light treatments. Dermatol Surg 26:835–43
- Caslini C, Connelly JA, Serna A et al. (2009) MLL associates with telomeres and regulates telomeric repeat-containing RNA transcription. Mol Cell Biol 29:4519–26
- Chang HY, Nuyten DS, Sneddon JB et al. (2005) Robustness, scalability and integration of a wound-response gene expression signature in predicting breast cancer survival. Proc Natl Acad Sci USA 102:3738–43
- Chen C, Liu Y, Liu Y et al. (2009) mTOR regulation and therapeutic rejuvenation of aging hematopoietic stem cells. Sci Signal 2:p.ra71
- Curran SP, Ruvkun G (2007) Lifespan regulation by evolutionarily conserved genes essential for viability. LoS Genet 3:e56
- ENCODE Consortium (2011) The user’s guide to the encyclopedia of DNA elements. PLoS Biol 9:1001046
- Fisher GJ, Varani J, Voorhees JJ (2008) Looking older: fibroblast collapse and therapeutic implications. Arch Dermatol 144:666–72
- Greer EL, Maures TJ, Hauswirth AG et al. (2010) Members of the histone H3 lysine 4 trimethylation complex regulate lifespan in germline-dependent manner in C. elegans. Nature 466:383–7
- Gupta RA, Shah N, Wang KC et al. (2010) Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 464: 1071–6
- Hsieh C-C, Kuro-o M, Rosenblatt KP et al. (2010) The ASK1-Signalosome regulates p38 MAPK activity in response to levels of endogenous oxidative stress in the Klotho mouse models of aging. Aging 2:597–611
- Hwang JS, Hwang JS, Chang I et al. (2007) Age-associated decrease in proteasome content and activities in human dermal fibroblasts: restoration of normal level of proteasome subunits reduces aging markers in fibroblasts from elderly persons. J Gerontol A Biol Sci Med Sci 62:490–9
- Itahana K, Zou Y, Itahana Y et al. (2003) Control of the replicative life span of human fibroblasts by p16 and the polycomb protein Bmi-1. Mol Cell Biol 23:389–401
- Kirby K, Hu J, Hilliker AJ et al. (2002) RNA interference-mediated silencing of Sod2 in Drosophila leads to early adult-onset mortality and elevated endogenous oxidative stress. Proc Natl Acad Sci USA 99:16162–7
- Langmead B, Trapnell C, Pop M et al. (2009) Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol 10:R25
- Lee Y, Kim H, Kim S et al. (2009) Activation of toll-like receptors 2, 3, or 5 induces matrix metalloproteinase-1 and -9 expression with the involvement of MAPKs and NFkB in human epidermal keratinocytes. Exp Dermatol 19:e44–9
- Liang R, Khanna A, Muthusamy S et al. (2011) Post-transcriptional regulation of IGF1R by key microRNAs in long-lived mutant mice. Aging cell 10:1080–8
- Liu L, Rando TA (2011) Manifestations and mechanisms of stem cell aging. J Cell Biol 103:257–66
- Loewer S, Cabili MN, Guttman M et al. (2010) Large intergenic non-coding RNA-RoR modulates reprogramming of human induced pluripotent stem cells. Nat Genet 42:1113–7
- Long X, Spycher C, Han ZS et al. (2002) TOR deficiency in C. elegans causes developmental arrest and intestinal atrophy by inhibition of mRNA translation. Curr Biol 12:1448–61
- de Magalhaes JP, Wuttke D, Wood SH et al. (2012) Genome-environment interactions that modulate aging: powerful targets for drug discovery. Pharmacol Rev 64:88–101
- Mayr C, Bartel DP (2009) Widespread shortening of 30UTRs by alternative cleavage and polyadenylation activates oncogenes in cancer cells. Cell 138:673–84
- McKenzie NE, Saboda K, Duckett LD et al. (2010) Development of A photographic scale for consistency and guidance in dermatologic assessment of forearm sun damage. Arch Dermatol 147:31–6
- Nelson TJ, Behfar A, Yamada S et al. (2009) Stem cell platforms for regenerative medicine. Clin Transl Sci 2:222–7
- Negishi K, Tezuka Y, Kushikata N et al. (2001) Photorejuvenation for Asian skin by intense pulsed light. Dermatol Surg 27:627–31
- Paddock HN, Schultz GS, Baker HV et al. (2003) Analysis of gene expression patterns in human postburn hypertrophic scars. J Burn Care Rehabil 24:371–7
- Parado J, Bougria M, Ayala A et al. (1999) Effects of aging on the various steps of protein synthesis: fragmentation of elongation factor 2. Free Radic Biol Med 26:362–70
- Partridge L (2010) The new biology of ageing. Phil Trans R Soc B 12 365:147–54
- Prieto VG, Sadick NS, Lloreta J et al. (2002) Effects of intense pulsed light on sun-damaged human skin, routine, and ultra-structural analysis. Lasers Surg Med 30:82–5
- Rodwell G, Sonu R, Zahn JM et al. (2004) A transcriptional profile of aging in the human kidney. PLoS Biol 2:e427
- Serrano M (2011) Cancer: final act of senescence. Nature 479:481–2
- Shapiro IM, Cheng AW, Flytzanis NC et al. (2011) An EMT-driven alternative splicing program occurs in human breast cancer and modulates cellular phenotype. PLoS Genet 7:e1002218
- Shaw T, Martin P (2009) Epigenetic reprogramming during wound healing: loss of polycomb-mediated silencing may enable upregulation of repair genes. EMBO Rep 10:881–6
- Tariq M, Kim HJ, Jejelowo O et al. (2011) Whole-transcriptome RNAseq analysis from minute amount of total RNA. Nucl Acids Res 39:e120
- Tazearslan C, Huang J, Barzilai N, Suh Y (2011) Impaired IGF1R signaling in cells expressing longevity-associated human IGF1R alleles. Aging cell 10:551–4
- Wang Z, Gersten M, Snyder M (2009) RNA-Seq: a revolutionary tool for transcriptomics. Nat Rev Genet 10:57–63
- Wapinski O, Chang HY (2011) Long noncoding RNAs and human disease. Trends Cell Biol 21:354–61
- Zahn JM, Sonu R, Vogel H et al. (2006) Transcriptional profiling of aging in human muscle reveals a common aging signature. PLoS Genet2:e115